




神经元膜的分子构造

厦门心理咨询-德仁心心理咨询机构:神经元膜的分子构造
神经解剖学和神经精神病学在整个脑和个别脑区水平上了解脑,神经生理学和神经 化学则从神经元水平研究脑。脑的主要功能是整合。正如整脑和部分脑区能进行整合机能那样,个别的单个神经元也能进行整合机能:既可接受和整合不同的传入信息,也可通过调制动作电位产生的频率及其末梢释放神经递质的种类与数量,对有关信息作出反应。大多数神经元从数以千百计的神经元接受突触输入,并且还受激素、免疫系统和机体时间节律性的影响。
神经元虽可视为脑的最小整合机制,但神经元自身的内部还存在重要的整合机制,如蛋白质机能的调节和基因表达的调控。神经元内的许多重要分子包括神经递质受体在内的蛋白质、酶、以及细胞骨架成分。由于蛋白质的机能是其形状和电荷的函数,那些能影响其形状和电荷的生化过程,如蛋白磷酸化,均可影响其机能。实际上,一个蛋白质分子的机能要受各种可逆的转录后修饰的调节;每一个转录后修饰均由突触输入、激素作用和时间生物学节律等不同来源的启动。
脑包括端脑、间脑、中脑、后脑、髓脑和脊髓等六个脑部。含有1014个神经元。神经元的种类千姿百态,至少有千余种以上。一个典型的神经元兩树突、胞体、轴突和轴突宋梢构成;从机能的角度,一个神经元哥以区分为输人、整合、传导和输出等相应的机能构造。输入部分将上一级输人神经元的化学信号转变为可分级的电信号;整合部分将分级电信号转变为频率偏码的全或无式的神经冲动或动作电位,并沿传导部分不衰减地传送到输出部分;输出部分,再依神经冲动的频率,释放相应数量的神经递质,将电信号转变为化学信号,传向下一级神经元。各级神经元的信息,沿着以“串联” 及/或“并联”方式连接起来的神经微环路,在脑内进行大范围的传递。所有这一系列信息传递活动均主要通过神经元膜来实现。
一、神经元膜的分子构造
神经元膜是神经细胞的重要组成部分,具有高度分化的分子构成和多种独特的生理功能。跨膜的物质转运和能量转换,神经元对细胞外物质的识别与结合、神经元跨膜信号传递与代谢调控,以及神经冲动的发生和扩布等生物学行为和过程,无一不与神经元 膜有关。
同其他细胞膜一样,神经元膜也很薄,在光镜下看不到,直到40年代后期才得以 用电镜进行观察和确认,1925年,Gorter和Grendel发现膜是一种具有两个分子厚度的脂肪性物质。10年后,Danielli和Davson认为,膜是由夹在两层蛋白质当中的双分子层 磷脂构成的“三夹板”,具有刚性排列顺序。1972年Singer和Nicolson提出的液态镶嵌模型(fluid mosaio model)看来是所有细胞膜的共同结构基础。液态镶嵌模型认为,膜的表面约有70%是脂质双分子层基质,其中“溶解”着既有亲水又有亲脂部分的兼性 (amphi pathic)蛋白质,进行受体、通道和泵等不同的功能活动(图2-1)。
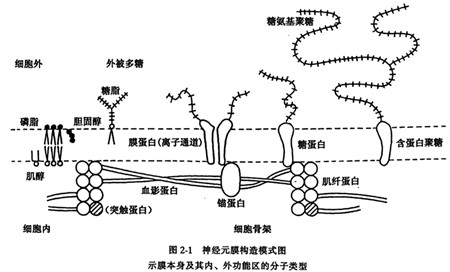
神经元膜的化学组成一般包括30%〜40%的蛋白质,40%〜50%的脂质以及1%〜 5%的糖,因膜的种类不同而有很大差异。通常膜的功能越是复杂,蛋白质所占的比例越大,反之则比例较低,仅起绝缘作用的神经髓鞘,仅含三种蛋白质,只占18%,脂质占70%,而功能复杂的线粒体内膜则含有60种蛋白质,占75%。
(一)脂质双层
脂质之间以及脂质与蛋白质之间的作用力主要是非共价键的静电、氢键、vander Waals相互作用。这些力的单独作用均比共价键弱,但这些力可以相加,产生相当稳定的联合。分子的解离基团和极性部分可与水分子的偶极相互作用而水化。如果分子与水的相互作用强于分子之间的相互作用,这种分子即是可溶的,大分子可能具有极化程度不同的表面功能区,其疏水部分可集合,形成微胞,最大程度地减少非极性表面对水的暴露,具有相互分隔的极化与非极性表面功能区的分子称为兼性分子,如去垢剂和复杂的脂质。
1.具有极性与非极性碱基的兼性分子易形成双层结构
磷脂是大多数细胞膜脂质的主要成分,通常形成脂质双层。每层包括甘油磷酸酯构成的极性头部和通常由两串酯化脂肪酸形成的碳氢化合物的非极性尾部。在水相环境中,头部与水和其他水相成分相互作用:而非极性的尾部主要是自身相互作用,形成单独的相,在水相环境中,大多数脂质因不同条件可以形成三种相结构,尽管在细胞膜只发现层状结构,但在某些膜转换过程中也可出现水包油或油包水状态的六角相。
2.脂质双层的物理状态具有多形性、流动性和不对称性等特征
含有磷脂的水相系统可表现不同的结构,呈固态晶体或液态晶体状态,纯磷脂可随温度变化,极易发生这两种相的转换。为维持细胞膜的正常机能活动,脂质双层须呈液晶态,磷脂分子可在双层平面内具有高度运动性,既可进行烃链围绕C—C键旋转而致的异构化活动,环绕与膜平面垂直的轴进行左右摆动和旋转运动,也可在膜内作侧向移动或侧向扩散以及在脂质双层中进行翻转运动。脂质分子在膜内侧的分布不是对称的,细胞膜内外侧面的成分因而有所不同:胞浆面含有较多的磷脂酰乙醇胺细胞外液面含有糖脂。脂质的这种不对称分布与膜蛋白的定向分布功能活动,膜两层上的电荷分布数量,流动性变动等均有密切关系。
(二)膜蛋白质
几乎所有的蛋白质均呈折叠状态,以使其主要的非极性氨基酸残基区域同其极性区 域相互分开。贯穿膜全长的膜整合蛋白质(integral protein)通常占膜蛋白的70%〜 80%,主要借疏水效应插入脂质双层。有时,膜的插人区只是作为一种锚,将在膜内侧 面起作用的蛋白质锚住。有些蛋白质几乎完全存在于脂质双层之间,可脱离锚区,在膜内变成完全主动的可溶蛋白质,分布于脂质双层表面的外周蛋白质通过静电力或van der Waals引力与膜结合,约占膜蛋白的20%〜30%。
1.插入膜的蛋白质节段几乎总是形成a螺旋
整合蛋白质的非极性氨基酸残基与脂质双层疏水核心相接触的部分,由于排除了水分子,多肽分子本身形成氢键的趋向大大增强,因而往往以a螺旋或P折叠,特别是a螺旋的形式存在。由于a螺旋的肽架形成一种氢键核心,螺旋节段的表面特性主要由氨 基酸侧链来决定。一个足以跨越脂质双层的a螺旋长度需要18—21个残基,只是为了 锚住蛋白质用的单个节段主要由疏水的氨基酸残基构成。
2.脂质双层的液态性质为整合蛋白质之间的相互作用提供条件
脂质双层的双向流动液态结构使蛋白质能在其中迅速扩散,进行垂直旋转和侧向移动,但不能进行从外层翻向内层或从内层翻向外层的翻转运动。某些神经递质和激素与其受体的相互作用可调制受体对换能器蛋白质的亲和力,进而影 响换能器蛋白质与效应器蛋白质的相互作用。这些相互作用均需在 膜双层中进行侧向扩散。细胞膜表面上的膜整合蛋白质的分布,可由于同外周蛋白质的特异相互作用而不同程度地受到约束。有些膜蛋白质通过细胞骨架蛋白质的网络分布于不同区域。
(三)神经元膜受体
受体必须与特异配体具有高度的亲和力,并以触发跨膜信号活动的构象变化对配体结合发生反应。细胞内固醇类激素受体也具备膜受体的这种双重特性,受体介导的跨膜信号活动的基本机制有三:①配体调节的离子通道。②配体调节的受体酶。③配体调节的受体一G蛋白激活:其结构基础分别与:①配体调节的寡聚体离子通道。②配体调 节的酪氨酸激酶。③G蛋白联结的视紫红质相关受体等三大受体族有关。
受体既见于突触后神经元,也见于突触前神经元。突触前受体的经典作用是作为突触前神经元的负反馈环路在起作用。例如,许多释放去甲肾上腺素的神经元有突触前电受体,如被其所释放的去甲肾上腺素占据,去甲肾4:腺素的释放即可减少或停止。与同一神经元自身的神经递质结合的突触前受体称为突触前同源受体;与不同神经元释放的神经递质结合的突触前受体称为突触前异源受体。
与受体有关的两个术语,“超敏性”和“低敏性”,分别指受体对衡量神经递质的反应分别高于和低于其通常的反应。受体的敏感性决定于受体的数目、受体对神经递质的亲和力、以及神经递质与受体结合借以转化为神经元内信使的效率。受体机能的这些步骤均是多变和可被调节的。
神经递质受体实际是心理治疗和精神活性药物的作用部位。分子生物学技术已鉴定出许多新的受体亚型及其序列,从而使有关病理学的研究更加精练,有关药物的设计更加准确。
1.与离子通道偶联的受体具有4〜5个跨膜功能区
与离子通道偶联的受体中,nAChR是第一个被纯化并用重组DNA技术阐明一级结构的受体,典型的nAChR是由电(3於等5个亚单位组成的。类似梅花瓣样的杂合五聚体,分子量接近280kD。每个亚单位由400余个氨基酸组成,共同围成一个对称轴心或中心腔,静息时关闭,激活时开放,直径达6.5人(0.65nm),允许Na+通透。
每个亚单位的肽链均有4〜5个跨膜功能区,称为M1~M4〜吣或M1,M2, M3,A,M4。每个跨膜节段均由疏水性很强的氨基酸组成,参与Na+通道形成。所有这些跨膜功能区均在200位残基之后出现。180〜200位氨基酸与激动剂和拮抗剂特异结合有关。128和145位半胱氨酸形成二硫键,形成一种发针式环。M2功能区的p和y链上的残基是铺在通道内里的序列,有丰富的苏氨酸和丝氨酸残基,使通道衬里呈亲水性,有助于离子流通过,其构象变化对通道启闭至关重要。
2.与G蛋白偶联的受体具有7个跨膜节段
与G蛋白偶联的受体族的成员与日俱增,已不下30余种,如毒蕈碱型乙酰胆碱受体。肾上腺素受体,多巴胺受体,5-羟色胺受体,P物质受体,K物质受体,神经肽类受体,组胺受体、腺苷受体等。
分子克隆结果显示这类受体也具有许多共同的特征。首先,这些受体的氨基酸排列顺序非常近似。其次,所有这些受体均有7个疏水区,形成Ⅰ~Ⅻ等7个跨膜的a-螺旋结构,每一跨膜区均由20〜25个疏水性很强的氨基酸组成,将受体嵌入膜内,再由亲水性氨基酸序列将它们联结起来。第三,这些受体与配体特,结合的部位不是在细胞外表面,而是陷人细胞膜内。第四,这类受体的N-末端较短,面向细胞外,上面有2个加糖基的部位;C-末端较长,伸入细胞内,有丰富的丝氨酸和苏氨酸残基,可供磷酸化。第五,这一受体家族的不同成员之间的差别主要在于细胞外的N-末端和细胞内的C-末端以及环状结构。最后,所有这些受体均通过与G蛋白的相互作用,实现信息的跨膜转导,其激活过程较慢(100〜250ms)。由于G蛋白性质的不同,受体与G蛋白相互作用的结果可导致腺苷酸环化酶抑制或兴奋、憐酸肌醇水解,或引起K+通道或其它离子通道的激活。一般认为n区的天冬氨酸参与配体结合;V〜竹跨膜节段的胞内亲水序列则可能与G蛋白结合。
3.与酪氨酸偶联的受体由单一肽链构成
这类受体亦称生长因子型神经肽受体,如神经生长因子受体、上皮生长因子受体、血小板生长因子受体以及胰岛素受体等。这些受体均由单一的肽链组成。受体的细胞外部分识别配体并与其特异结合;受体的细胞内部分则具有激酶活性,使受体自身的酪氨 酸残基磷酸化,从而发挥其特定的调节作用。
尊重您的个人私密,使隐私权得到充分的保障。
关注心理健康(预约) 0592—5515516
官方网站-心理健康:http://www.xmxljg.com
专家微博-心理语录:http://weibo.com/fjxlzx
专家博客-心理案例:http://blog.sina.com.cn/xmxlys
企业微博-心理视频:http://e.weibo.com/xmxljg
企业博客-心理课堂:http://blog.sina.com.cn/xmxljg
↓
权威心理督导:郭潇赢是著名心理学教授/国家级心理治疗师
厦门心理咨询-德仁心心理咨询机构(尊重您的个人私密,使隐私权得到充分的保障)
